近日,4166金沙之选主页通道、海洋与地球学院海洋计算生物地球化学研究组在mSystems期刊上以“N2 fixation in Trichodesmium does not require spatial segregation from photosynthesis”为题发表研究论文。该研究通过束毛藻生态生理模型实验,探索了束毛藻光合和固氮作用的共存机制,为理解海洋生物固氮过程与机制提供了新视角。
生物固氮为海洋贡献了高达一半的生物可利用氮。在总体缺氮的表层海洋中,以自养蓝藻为主要种群完成的固氮作用,是支持初级生产力的关键过程之一。
然而,自养蓝藻进行光合作用产生的氧气,对固氮过程有很强的抑制作用。海洋固氮蓝藻通过形成具有厚壁的异形胞以有效隔绝氧气,或在无光合作用的夜间时段进行固氮。但海洋中的优势固氮类群——束毛藻,却很特殊,其不形成异形胞,却能在白天时段同时进行光合和固氮作用。那么,束毛藻是如何避免光合作用产生的氧气对固氮过程的抑制作用呢?
针对这个问题,学者们主要提出时间错峰和空间隔离两种假说。时间错峰假说提出,光合和固氮作用在日周期中达到峰值的时间不同(图1A),可能有利于在固氮时维持低胞内氧气浓度。然而,大量观测数据显示,在固氮峰值期仍有较高水平的光合作用和氧气产生速率。那么,仅通过光合和固氮作用的错峰,束毛藻是否足以解决光合产氧和固氮之间的矛盾呢?部分主流科学家基于免疫荧光法的证据,提出空间隔离的假说,即在束毛藻藻丝或者束状体中,各段细胞分别进行光合和固氮作用,从而形成二者空间上的隔离(图1B)。然而,近年来通过更先进的研究方法(如NanoSIMS),学者并未在束毛藻中发现光合和固氮作用的空间隔离现象。那么,束毛藻光合和固氮作用的空间隔离,是否必须存在呢?
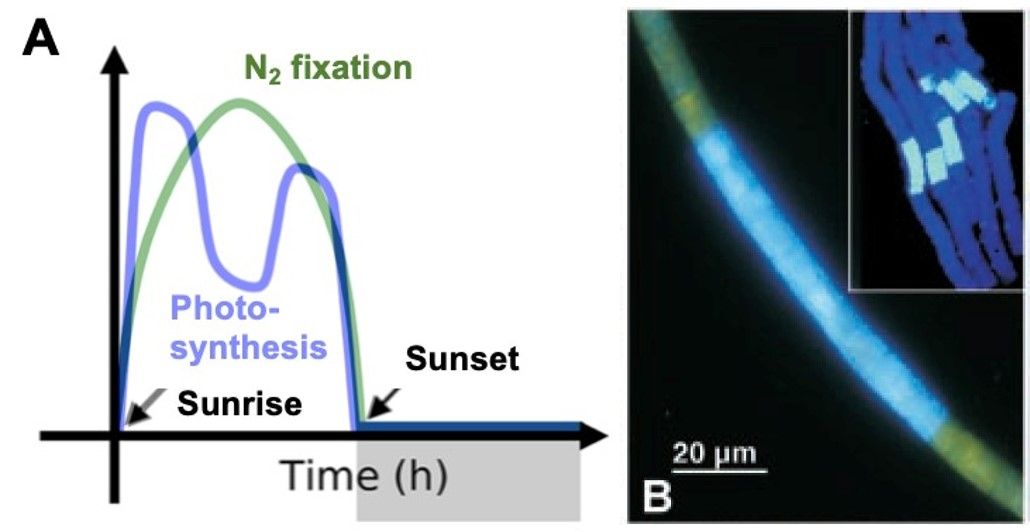
图1. (A)束毛藻光合和固氮速率日间变化示意图(改自Inomura et al. 2019, mSystems);(B)免疫荧光法显示束毛藻固氮酶集中分布于一段细胞(来自Berman-Frank et al. 2001, Science)
本研究通过构建束毛藻细胞生态生理模型(图2),模拟了其进行光合和固氮作用的主要过程,细胞内氧气的产生、消耗和交换,以及胞内能量的产生和分配,较好地还原了历史实验室数据中束毛藻光合和固氮速率的日间变化特征,进一步提出束毛藻不依赖于空间隔离的光合和固氮作用的共存机制(图3):1)上午时段,高速率的光合作用积累了大量有机碳,期间产生的氧气逐步扩散出细胞。2)中午到下午时段,虽然光合作用和产氧速率仍维持在中等水平,但束毛藻通过大量呼吸上午积累的有机碳,消耗胞内氧气(称之为“呼吸保护”);同时,由于细胞膜对氧气的低透过性,氧气难以从胞外向胞内扩散。这一关键机制使胞内氧气浓度降到极低水平,为固氮形成低氧窗口。模型实验结果显示,呼吸保护消耗了大量的有机碳,造成光合固碳和固氮的高摩尔比,这与大量实验室数据(约30:1 – 50:1)一致。
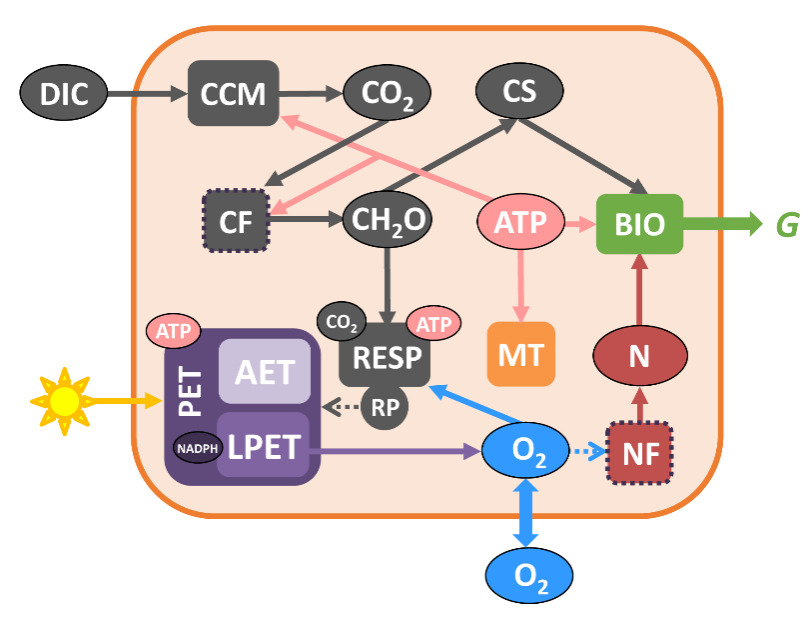
图2. 束毛藻细胞生态生理模型示意图
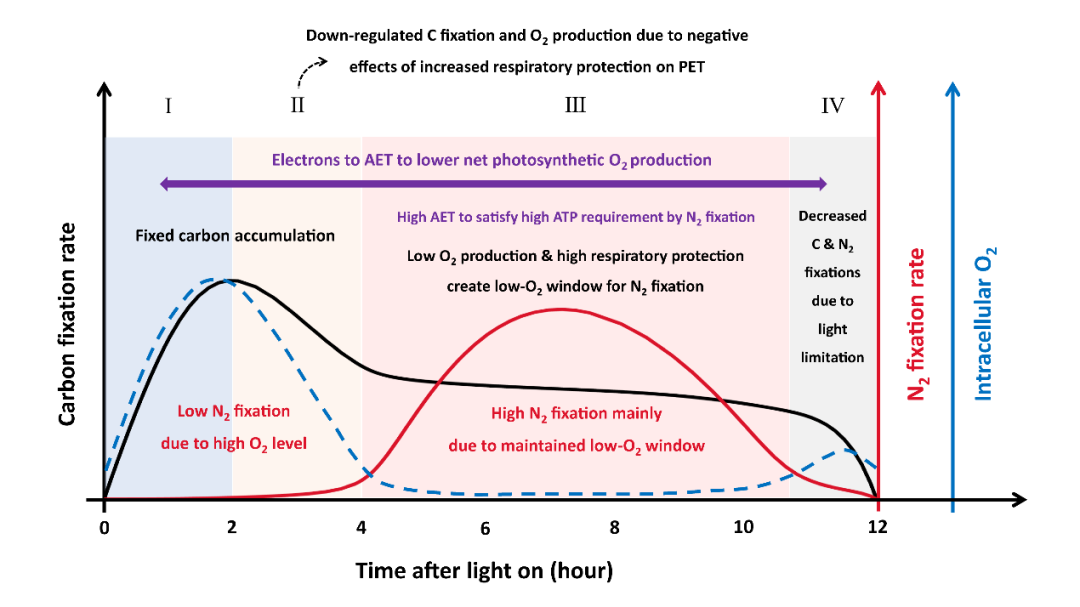
图3. 模型提出的束毛藻日间光合作用和固氮作用的共存机制
该研究通过进一步的模型实验,评估了光合和固氮作用的空间隔离对束毛藻生长速率的影响。研究人员在模型中分配不同细胞(光合细胞和固氮细胞)分别进行光合作用和固氮作用。假设所合成的碳、氮可以无损耗地在细胞间高效输送,则束毛藻的生长速率将提高1倍(图3)。但是,目前在束毛藻的细胞间未发现氮转运通道。如果固氮细胞固定的氮必须先释放到环境中才能被光合细胞吸收,则氮在细胞间的转运必然导致损耗。模型实验结果显示,当细胞间氮输送的损耗比例较高时,空间隔离不会显著促进(甚至会降低)束毛藻的生长速率(图4)。
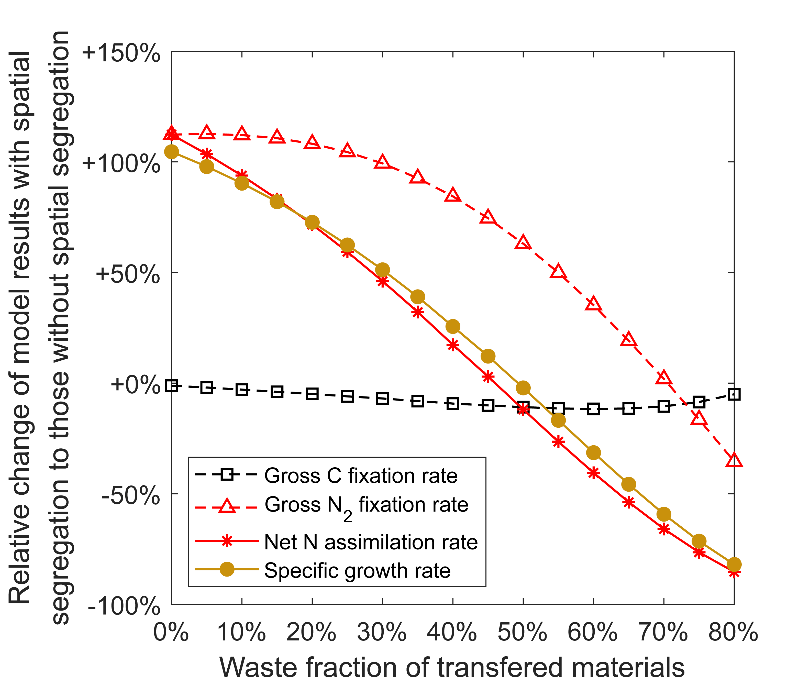
图4. 光合和固氮作用空间隔离下,模型实验中束毛藻固氮和生长速率与物质转运损耗比例的关系
本研究结合已有理论和实验室数据,构建了束毛藻生态生理模型,对海洋固氮生物如何有效调控胞内氧气以利于固氮进行了定量分析,得出以下观点:1)束毛藻通过日间时段光合和固氮作用在时间上的错峰,即可在单个细胞内解决光合产氧对固氮的抑制这一难题;2)束毛藻的光合和固氮作用可能不存在空间隔离。
该论文第一作者为4166金沙之选主页通道博士生罗伟成,通讯作者为罗亚威副教授,共同作者还包括美国罗德岛大学Keisuke Inomura助理教授和山东大学博士生张涵。本研究获得国家自然科学基金面上项目(42076153)和重大项目Carbon-FE(41890802)的联合资助。
论文链接
https://journals.asm.org/doi/10.1128/msystems.00538-22

